

A network of assembly factors is involved in remodeling rRNA elements during preribosome maturation
- PMID: 25404745
- PMCID: PMC4242840
- DOI: 10.1083/jcb.201408111
A network of assembly factors is involved in remodeling rRNA elements during preribosome maturation
Erratum in
-
A network of assembly factors is involved in remodeling rRNA elements during preribosome maturation.J Cell Biol. 2015 Jul 6;210(1):169-70. doi: 10.1083/jcb.20140811106112015c. J Cell Biol. 2015. PMID: 26150393 Free PMC article. No abstract available.
Abstract
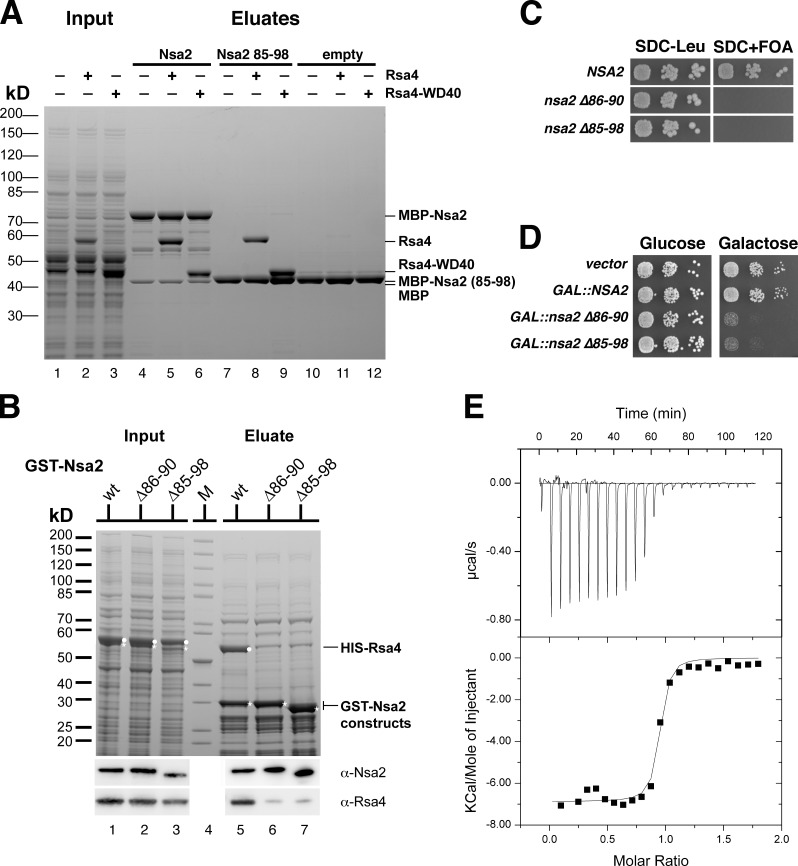
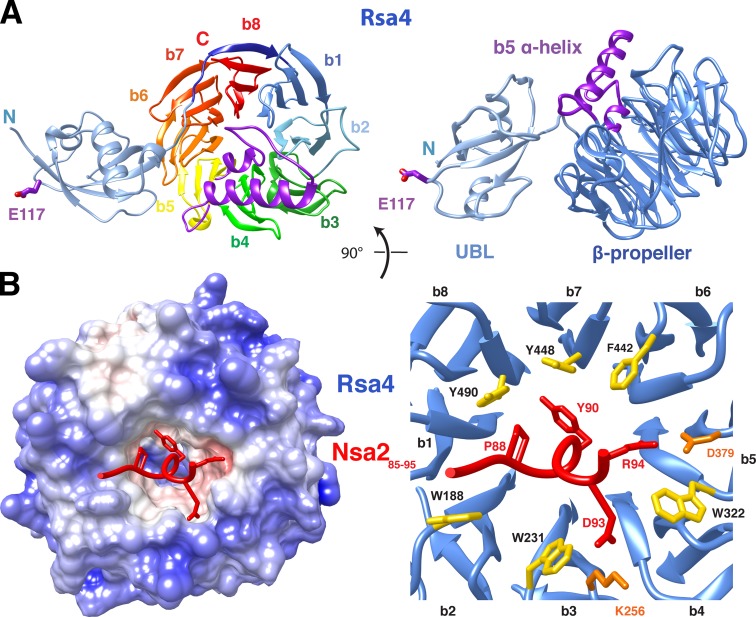
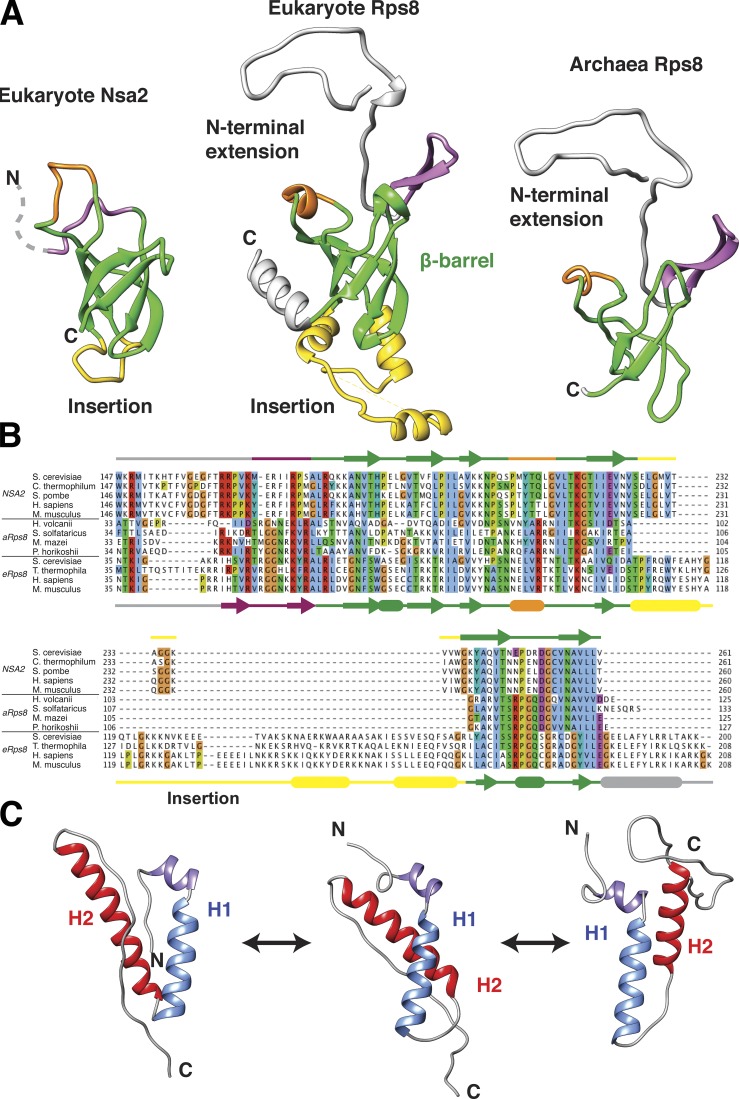
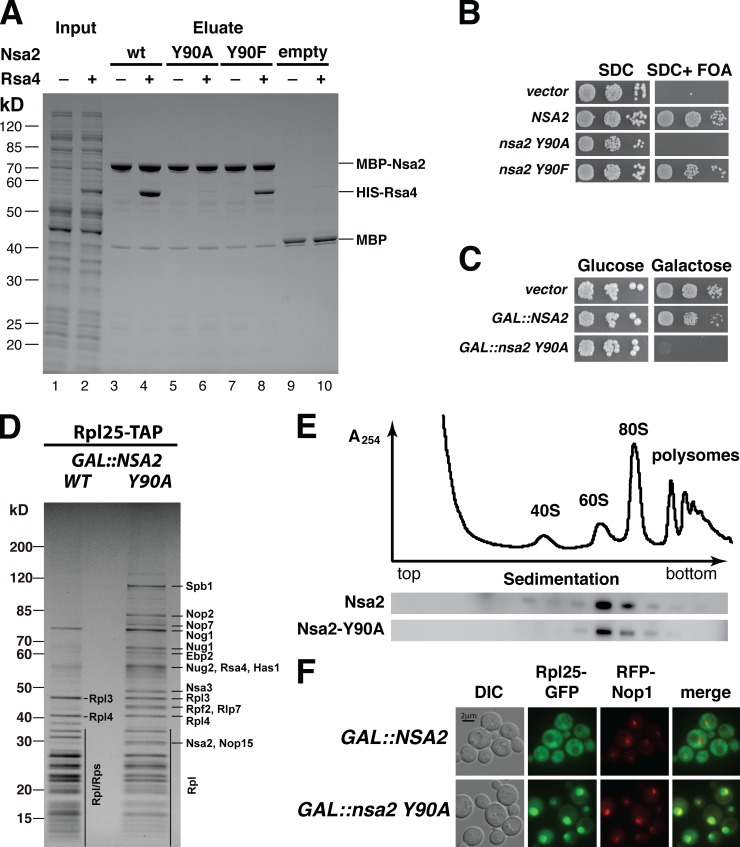
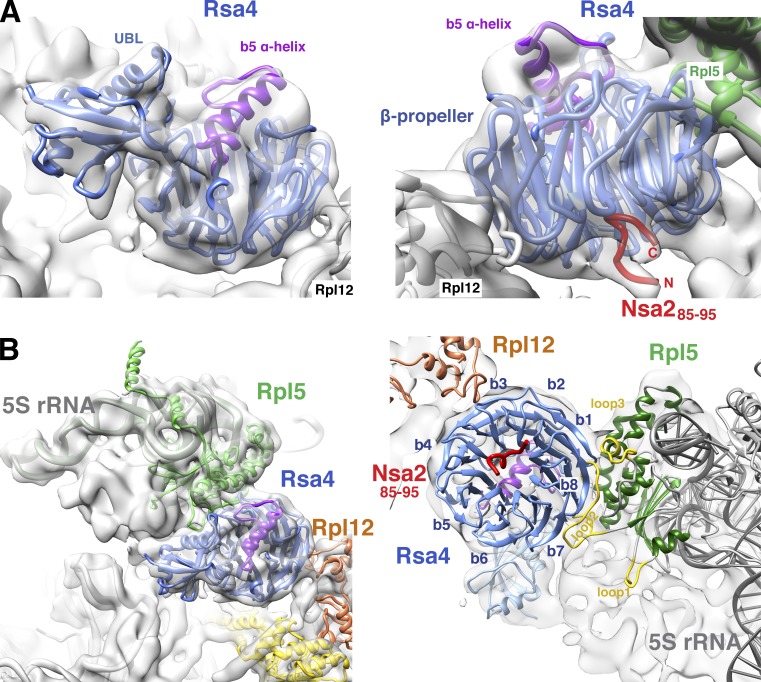
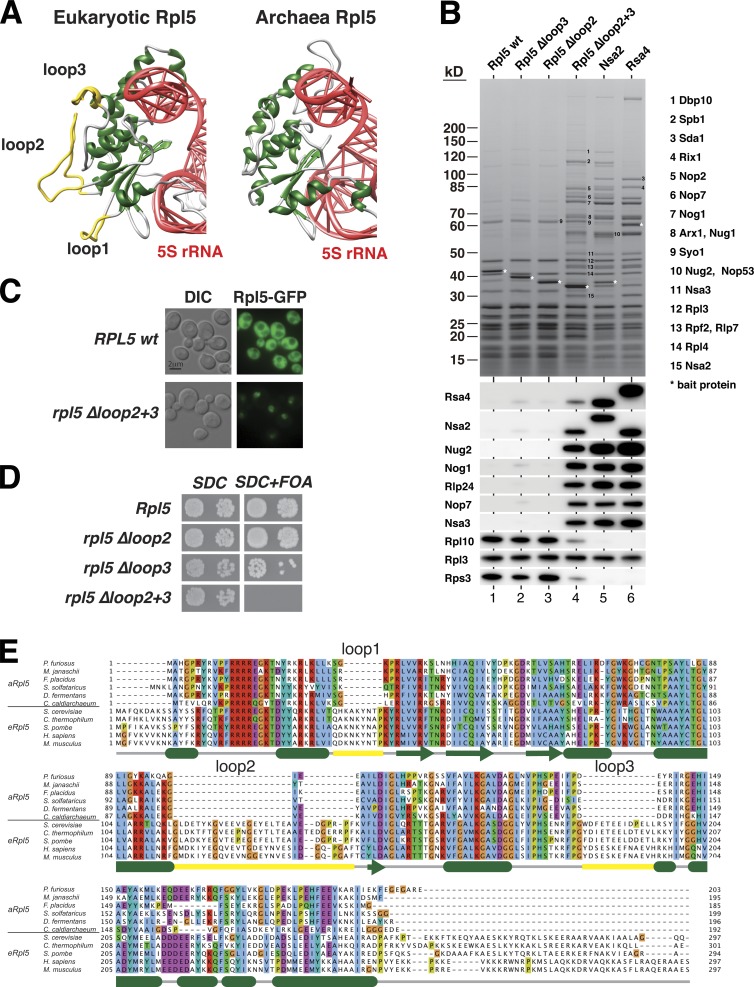
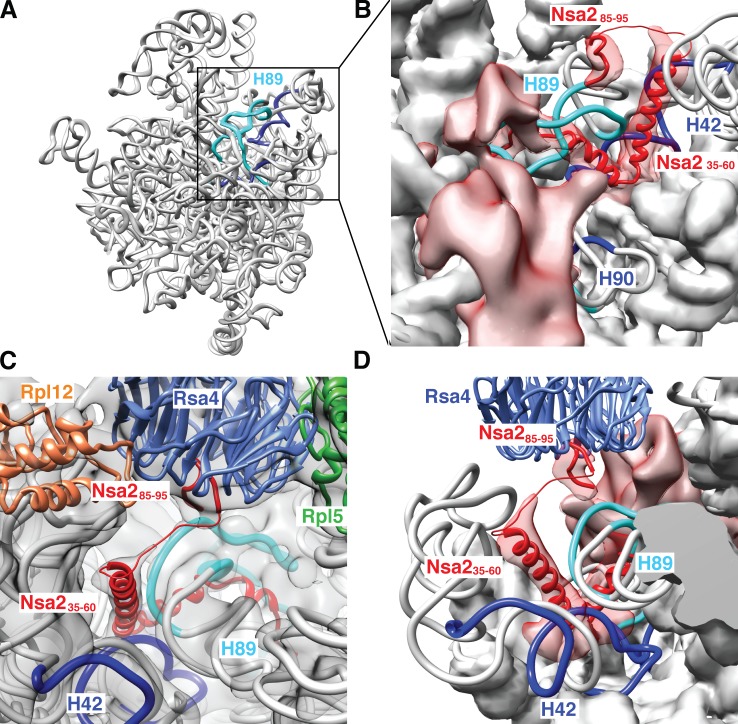
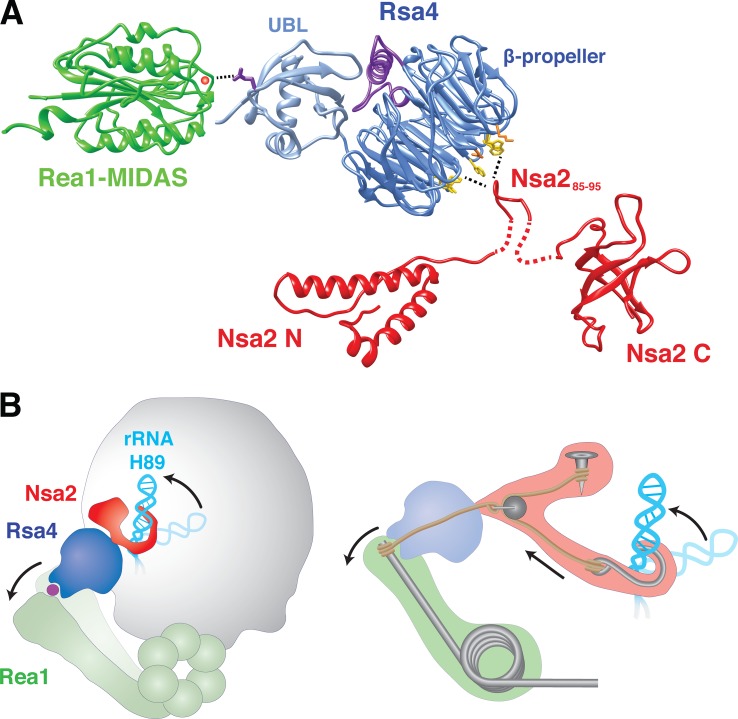
Eukaryotic ribosome biogenesis involves ∼200 assembly factors, but how these contribute to ribosome maturation is poorly understood. Here, we identify a network of factors on the nascent 60S subunit that actively remodels preribosome structure. At its hub is Rsa4, a direct substrate of the force-generating ATPase Rea1. We show that Rsa4 is connected to the central protuberance by binding to Rpl5 and to ribosomal RNA (rRNA) helix 89 of the nascent peptidyl transferase center (PTC) through Nsa2. Importantly, Nsa2 binds to helix 89 before relocation of helix 89 to the PTC. Structure-based mutations of these factors reveal the functional importance of their interactions for ribosome assembly. Thus, Rsa4 is held tightly in the preribosome and can serve as a "distribution box," transmitting remodeling energy from Rea1 into the developing ribosome. We suggest that a relay-like factor network coupled to a mechano-enzyme is strategically positioned to relocate rRNA elements during ribosome maturation.
© 2014 Baßler et al.
Figures
References
-
- Adams P.D., Afonine P.V., Bunkóczi G., Chen V.B., Davis I.W., Echols N., Headd J.J., Hung L.W., Kapral G.J., Grosse-Kunstleve R.W., et al. . 2010. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 66:213–221 10.1107/S0907444909052925 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
