

Allostery and substrate channeling in the tryptophan synthase bienzyme complex: evidence for two subunit conformations and four quaternary states
- PMID: 23952479
- PMCID: PMC4254854
- DOI: 10.1021/bi400795e
Allostery and substrate channeling in the tryptophan synthase bienzyme complex: evidence for two subunit conformations and four quaternary states
Abstract
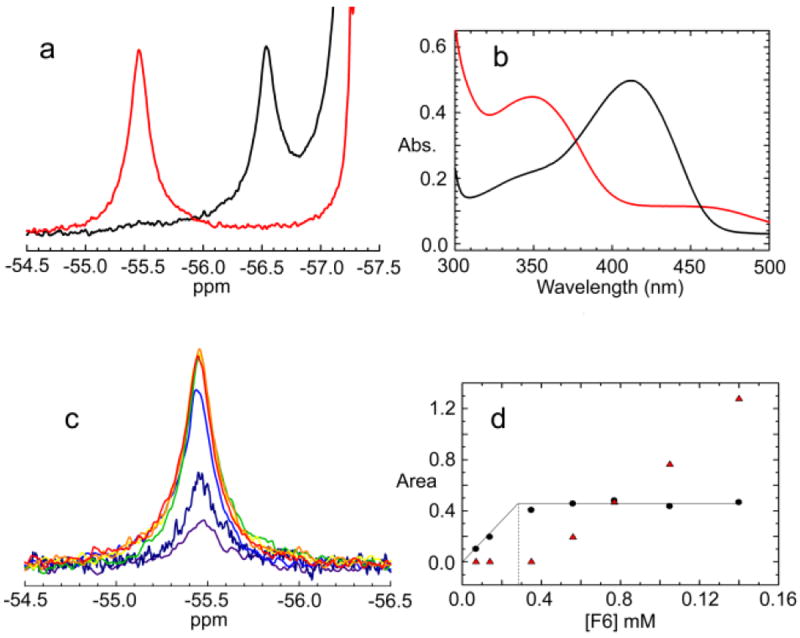
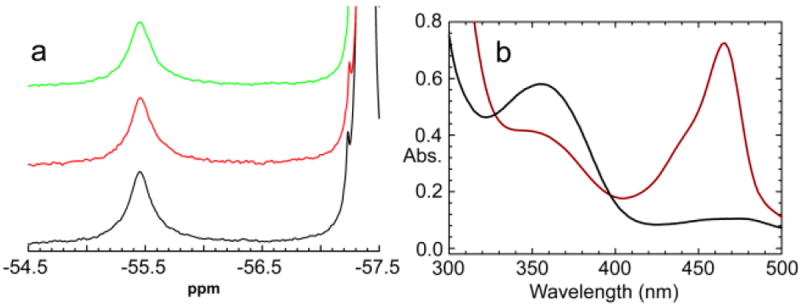
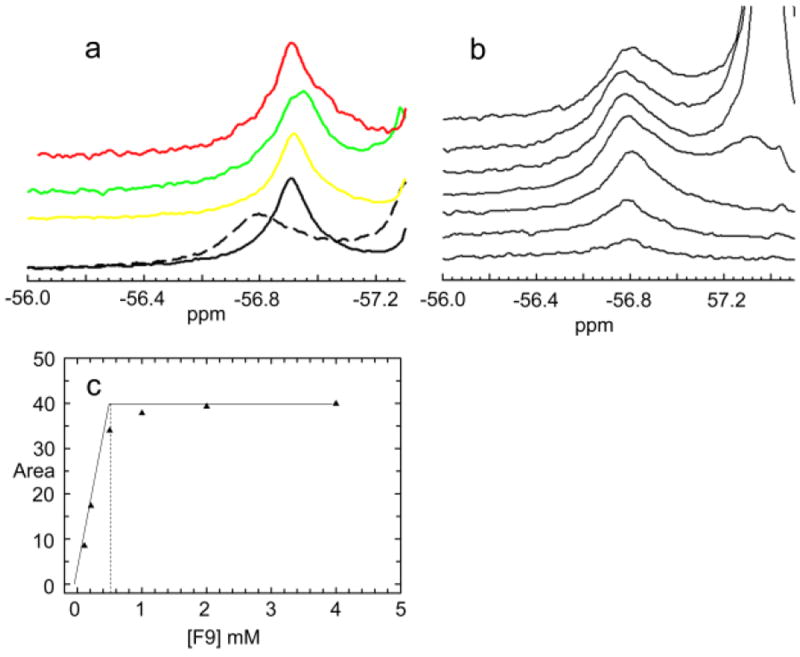
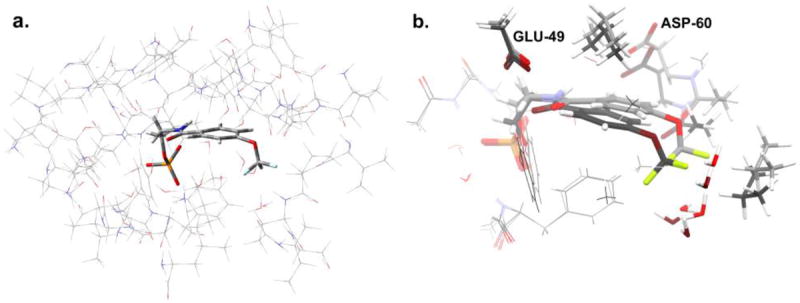
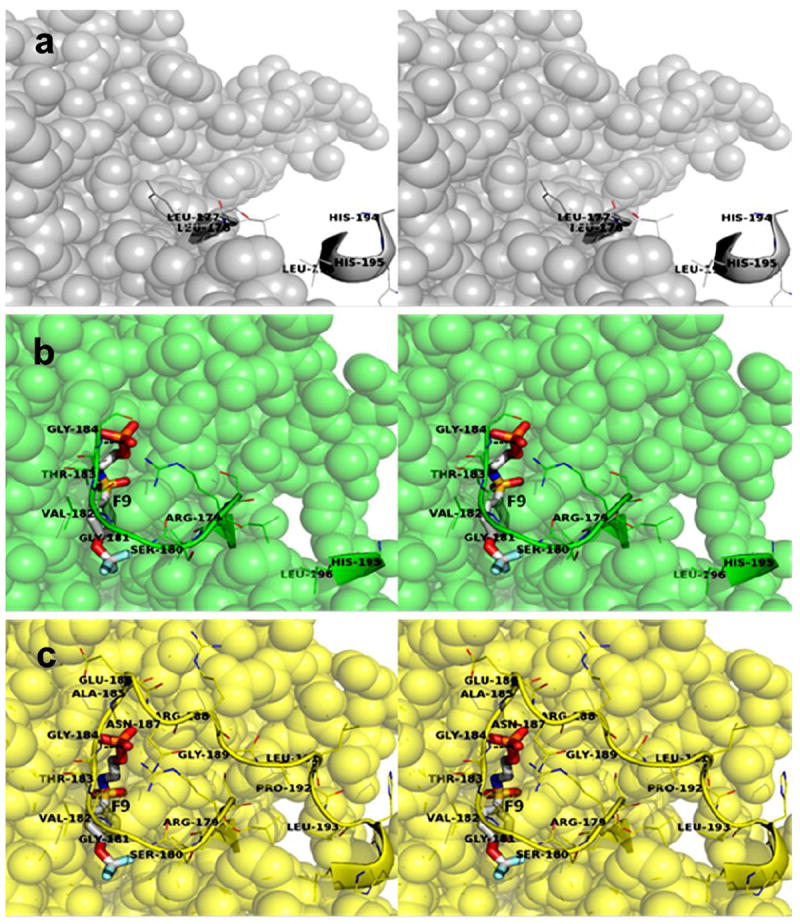
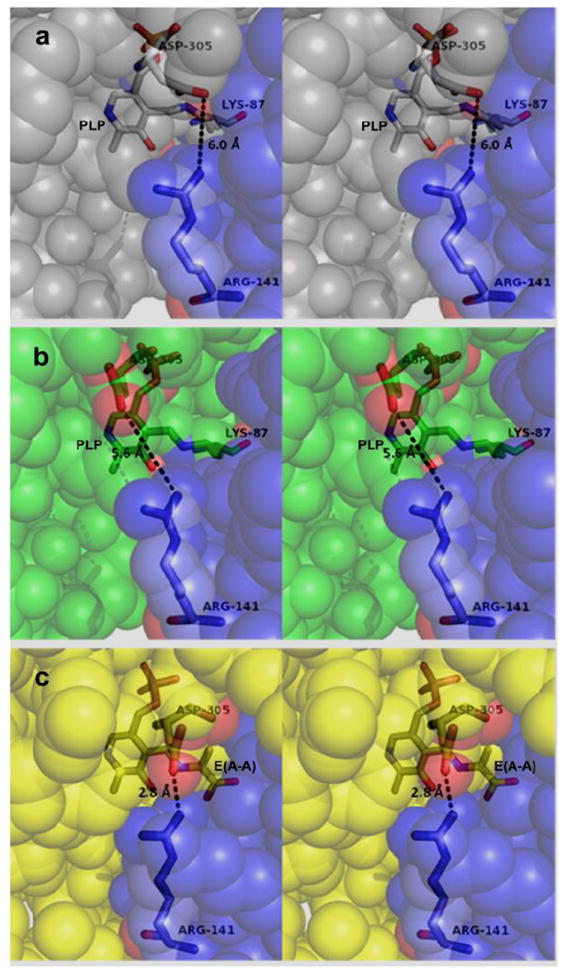
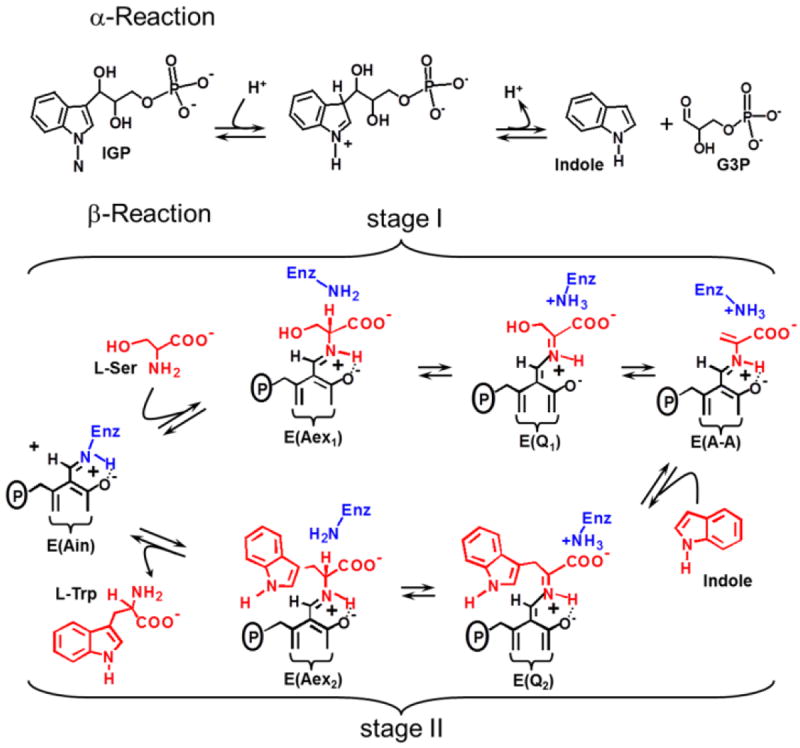
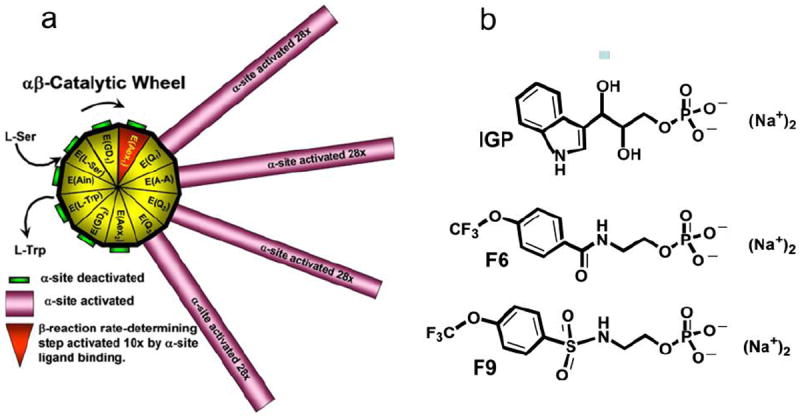
The allosteric regulation of substrate channeling in tryptophan synthase involves ligand-mediated allosteric signaling that switches the α- and β-subunits between open (low activity) and closed (high activity) conformations. This switching prevents the escape of the common intermediate, indole, and synchronizes the α- and β-catalytic cycles. (19)F NMR studies of bound α-site substrate analogues, N-(4'-trifluoromethoxybenzoyl)-2-aminoethyl phosphate (F6) and N-(4'-trifluoromethoxybenzenesulfonyl)-2-aminoethyl phosphate (F9), were found to be sensitive NMR probes of β-subunit conformation. Both the internal and external aldimine F6 complexes gave a single bound peak at the same chemical shift, while α-aminoacrylate and quinonoid F6 complexes all gave a different bound peak shifted by +1.07 ppm. The F9 complexes exhibited similar behavior, but with a corresponding shift of -0.12 ppm. X-ray crystal structures show the F6 and F9 CF3 groups located at the α-β subunit interface and report changes in both the ligand conformation and the surrounding protein microenvironment. Ab initio computational modeling suggests that the change in (19)F chemical shift results primarily from changes in the α-site ligand conformation. Structures of α-aminoacrylate F6 and F9 complexes and quinonoid F6 and F9 complexes show the α- and β-subunits have closed conformations wherein access of ligands into the α- and β-sites from solution is blocked. Internal and external aldimine structures show the α- and β-subunits with closed and open global conformations, respectively. These results establish that β-subunits exist in two global conformational states, designated open, where the β-sites are freely accessible to substrates, and closed, where the β-site portal into solution is blocked. Switching between these conformations is critically important for the αβ-catalytic cycle.
Conflict of interest statement
The authors have no competing interests.
Figures

References
-
- Monod J, Wyman J, Changeux JP. On the nature of allosteric transitions: a plausible model. J Mol Biol. 1965;12:88–118. - PubMed
-
- Grant G. Allosteric regulation. Arch Biochem Biophys. 2012;519:67–231. Guest editor. - PubMed
-
- Batiza AF, Rayment I, Kung C. Channel gate! Tension, leak and disclosure. Structure. 1999;7:99–103. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
