

The aspartate-less receiver (ALR) domains: distribution, structure and function
- PMID: 25875291
- PMCID: PMC4395418
- DOI: 10.1371/journal.ppat.1004795
The aspartate-less receiver (ALR) domains: distribution, structure and function
Erratum in
-
Correction: The Aspartate-Less Receiver (ALR) Domains: Distribution, Structure and Function.PLoS Pathog. 2015 Jun 18;11(6):e1004951. doi: 10.1371/journal.ppat.1004951. eCollection 2015 Jun. PLoS Pathog. 2015. PMID: 26086524 Free PMC article. No abstract available.
Abstract
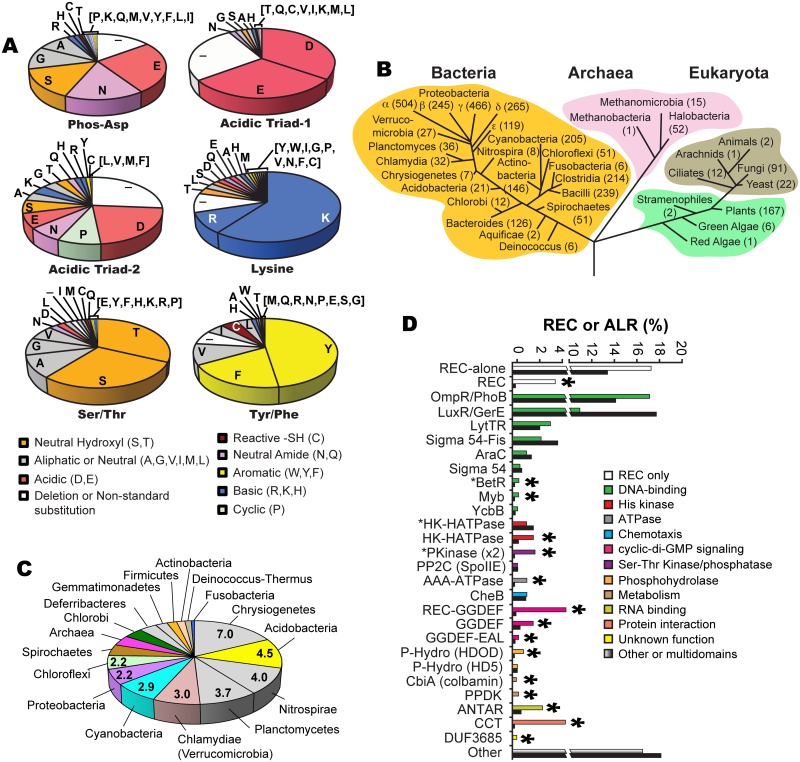
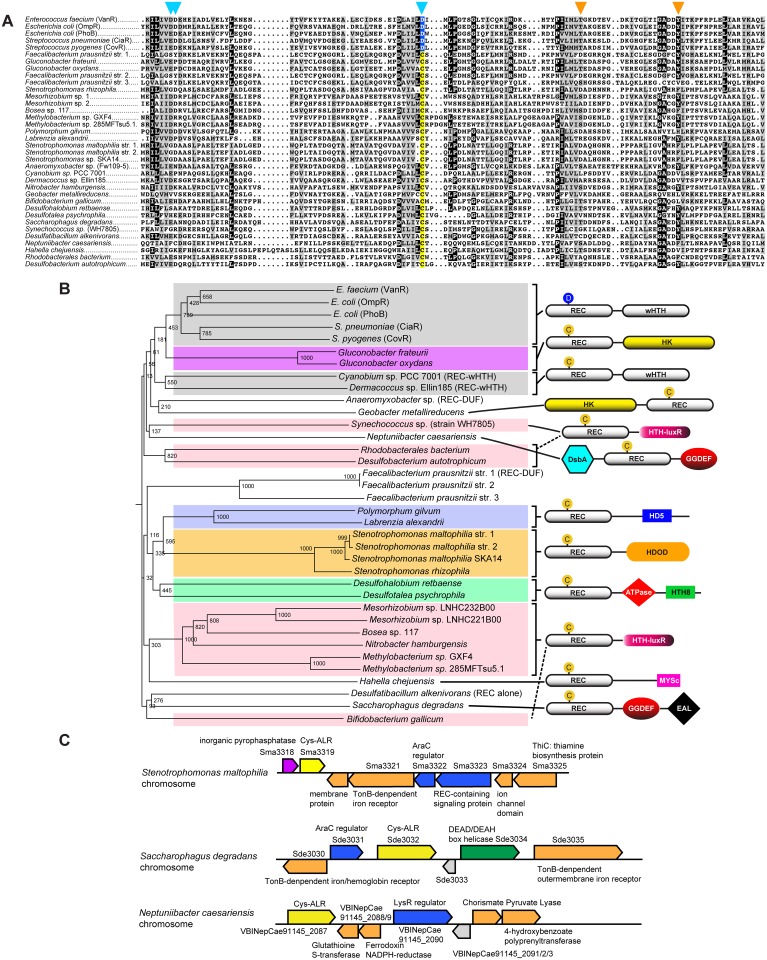
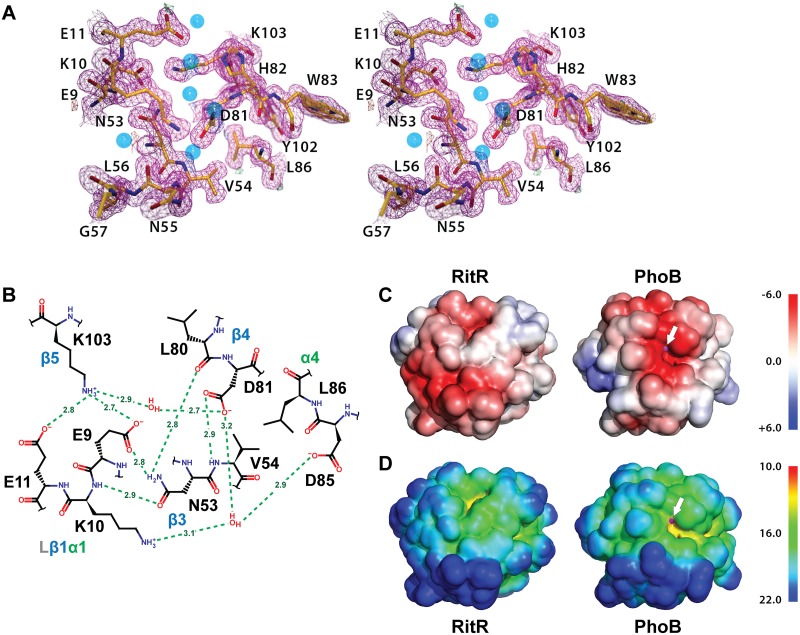
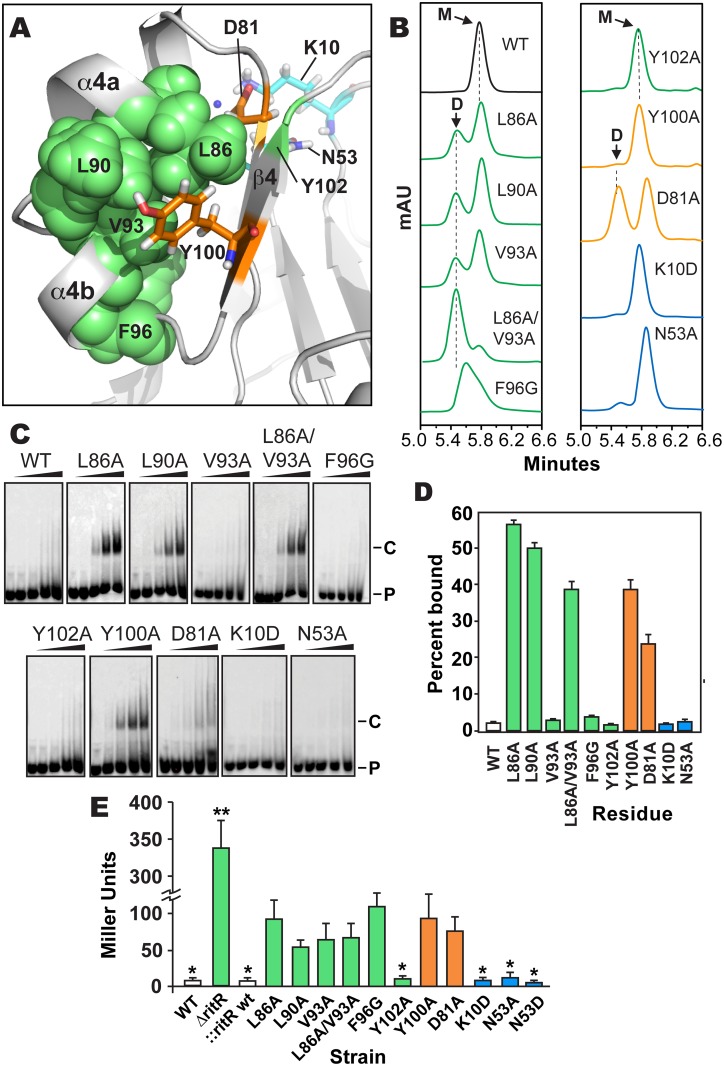
Two-component signaling systems are ubiquitous in bacteria, Archaea and plants and play important roles in sensing and responding to environmental stimuli. To propagate a signaling response the typical system employs a sensory histidine kinase that phosphorylates a Receiver (REC) domain on a conserved aspartate (Asp) residue. Although it is known that some REC domains are missing this Asp residue, it remains unclear as to how many of these divergent REC domains exist, what their functional roles are and how they are regulated in the absence of the conserved Asp. Here we have compiled all deposited REC domains missing their phosphorylatable Asp residue, renamed here as the Aspartate-Less Receiver (ALR) domains. Our data show that ALRs are surprisingly common and are enriched for when attached to more rare effector outputs. Analysis of our informatics and the available ALR atomic structures, combined with structural, biochemical and genetic data of the ALR archetype RitR from Streptococcus pneumoniae presented here suggest that ALRs have reorganized their active pockets to instead take on a constitutive regulatory role or accommodate input signals other than Asp phosphorylation, while largely retaining the canonical post-phosphorylation mechanisms and dimeric interface. This work defines ALRs as an atypical REC subclass and provides insights into shared mechanisms of activation between ALR and REC domains.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

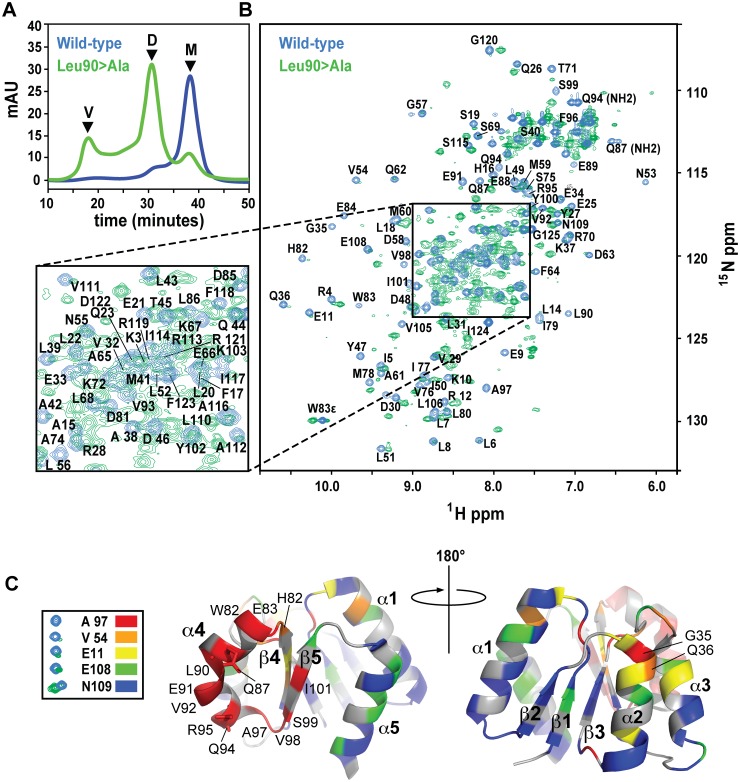
References
-
- Hoch JA, Silhavy TJ, editors (1995) Two-component signal transduction. Washington, D.C.: American Society for Microbiology.
-
- Volkman BF, Lipson D, Wemmer DE, Kern D (2001) Two-state allosteric behavior in a single-domain signaling protein. Science 291: 2429–2433. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
