

A subfamily roadmap of the evolutionarily diverse glycoside hydrolase family 16 (GH16)
- PMID: 31501245
- PMCID: PMC6827312
- DOI: 10.1074/jbc.RA119.010619
A subfamily roadmap of the evolutionarily diverse glycoside hydrolase family 16 (GH16)
Abstract
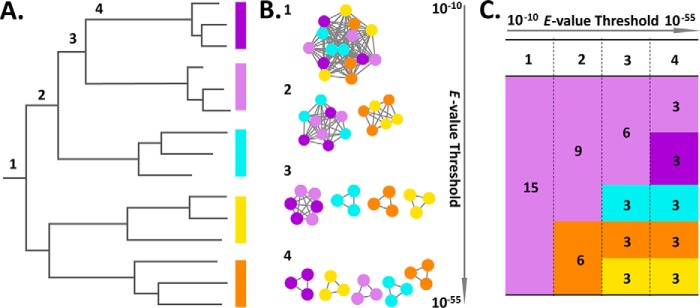
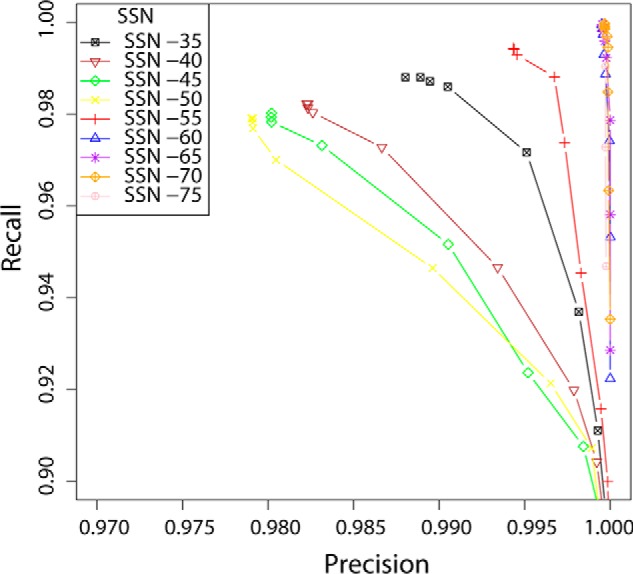
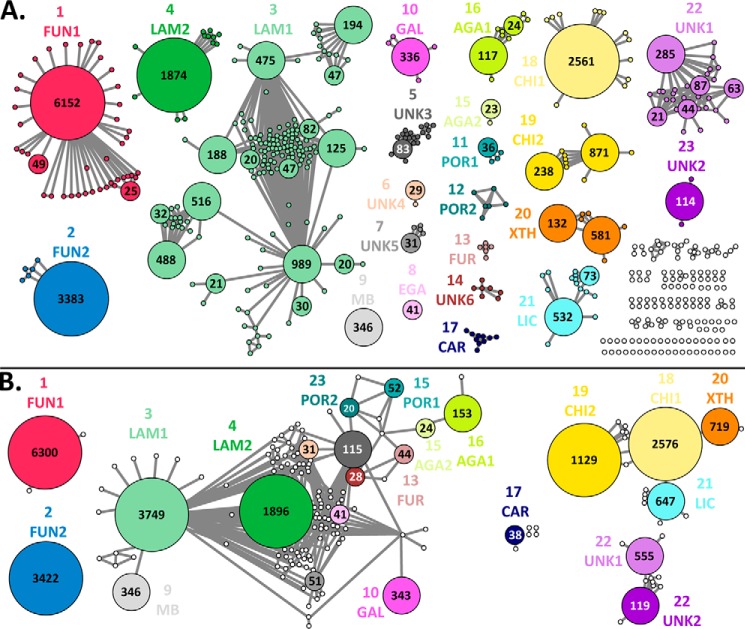
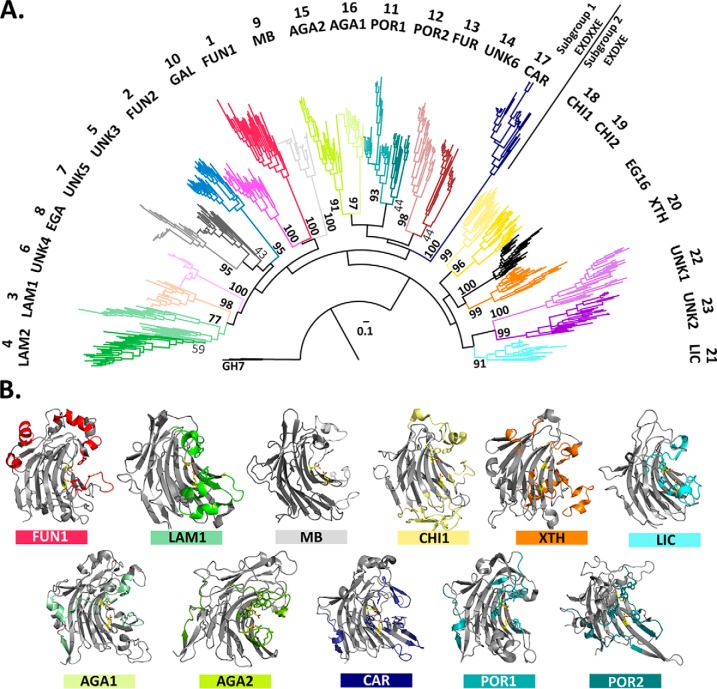
Glycoside hydrolase family (GH) 16 comprises a large and taxonomically diverse family of glycosidases and transglycosidases that adopt a common β-jelly-roll fold and are active on a range of terrestrial and marine polysaccharides. Presently, broadly insightful sequence-function correlations in GH16 are hindered by a lack of a systematic subfamily structure. To fill this gap, we have used a highly scalable protein sequence similarity network analysis to delineate nearly 23,000 GH16 sequences into 23 robust subfamilies, which are strongly supported by hidden Markov model and maximum likelihood molecular phylogenetic analyses. Subsequent evaluation of over 40 experimental three-dimensional structures has highlighted key tertiary structural differences, predominantly manifested in active-site loops, that dictate substrate specificity across the GH16 evolutionary landscape. As for other large GH families (i.e. GH5, GH13, and GH43), this new subfamily classification provides a roadmap for functional glycogenomics that will guide future bioinformatics and experimental structure-function analyses. The GH16 subfamily classification is publicly available in the CAZy database. The sequence similarity network workflow used here, SSNpipe, is freely available from GitHub.
Keywords: Hidden Markov Model (HMM); beta-jelly-roll fold; beta-sandwich; carbohydrate-active enzymes (CAZymes); enzyme structure; glycoside hydrolase; phylogenetics; protein evolution; sequence similarity networks (SSN); structural biology.
© 2019 Viborg et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures

References
-
- Varki A. (2017) Essentials of Glycobiology, 3rd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
